
研究表明,大部份鈣粘素蛋白家族與疾病,如腫瘤的發(fā)生相關(guān)。由于細(xì)胞之間的黏附需要鈣粘素蛋白同源二聚化,相應(yīng)的,能夠調(diào)節(jié)二聚化過(guò)程的小分子,將可能成為潛在的治療性藥物。因此,本文介紹了P-鈣粘素特異性小分子的鑒定,作用于P-鈣粘素蛋白介導(dǎo)的細(xì)胞黏附過(guò)程。雖然該小分子是一種片段化的化合物,但經(jīng)實(shí)驗(yàn)證明,它可以結(jié)合到P-鈣粘素蛋白的凹陷區(qū),此前該位點(diǎn)還未做為藥物靶點(diǎn)。該分子片段能夠阻止一種被稱為X二聚物的中間體形成中的關(guān)鍵氫鍵的形成,進(jìn)而調(diào)節(jié)了X二聚化的過(guò)程。我們的發(fā)現(xiàn)將對(duì)小分子用于蛋白-蛋白之間的相互作用調(diào)節(jié)以及蛋白復(fù)合物組裝策略產(chǎn)生重要的影響。
蛋白的組裝揭示了幾乎所有生物學(xué)過(guò)程:1. 已有大量被報(bào)道的各種功能復(fù)合物的同源或異源結(jié)構(gòu)信息;2. 能夠?qū)Φ鞍捉M裝進(jìn)行調(diào)節(jié)的調(diào)節(jié)分子可以做為潛在的藥物或探針,用于進(jìn)一步研究蛋白的功能。但小分子在調(diào)節(jié)蛋白-蛋白相互作用及組裝方面存在困難,部分原因如下:首先,蛋白互作通常會(huì)牽涉到較大的分子區(qū)域,但是,小分子可以進(jìn)入的區(qū)域較小。除此之外,在蛋白-蛋白互作界面,通常不存在明顯的凹槽;缺少能夠指導(dǎo)配體分子設(shè)計(jì)的天然蛋白互作分子抑制劑存在。細(xì)胞通過(guò)鈣粘素蛋白家族,一類鈣依賴性的細(xì)胞黏附蛋白分子,發(fā)生細(xì)胞之間的黏附,蛋白的組裝。根據(jù)不同的生物組織環(huán)境,鈣粘素分子在各種組織的腫瘤發(fā)生中發(fā)揮作用。P-鈣粘素分子是一種經(jīng)典的鈣粘蛋白,已發(fā)現(xiàn)在許多腫瘤組織中過(guò)表達(dá),促進(jìn)了腫瘤的轉(zhuǎn)移和增殖。細(xì)胞之間的聚集體形成,能夠阻止anoikis和P-鈣粘素蛋白介導(dǎo)的信號(hào)通路,這對(duì)于腫瘤細(xì)胞的存活至關(guān)重要。因此,P-鈣粘素蛋白功能的抑制已成為一種潛在的抗癌策略。細(xì)胞黏附可以通過(guò)不同的細(xì)胞間的鈣粘素蛋白反式同源二聚化實(shí)現(xiàn),也可以通過(guò)相同細(xì)胞間的鈣粘素蛋白的順式聚集而成。對(duì)于一些類型I的經(jīng)典的鈣粘素蛋白,包括P-鈣粘素,反式同源二聚化已經(jīng)被廣泛的研究。蛋白五個(gè)結(jié)構(gòu)域中的兩個(gè)細(xì)胞外結(jié)構(gòu)域(EC12)經(jīng)證實(shí)在蛋白互作的過(guò)程中發(fā)揮作用(請(qǐng)見(jiàn)Fig. 1a)。X二聚物,也被稱為鏈-插入二聚物(S-S dimer),是一種中間過(guò)渡狀態(tài),能夠促進(jìn)最終二聚體的形成。S-S二聚物具有一種特定的結(jié)合模式,在該結(jié)合模式之下,一個(gè)單體N末端的色氨酸殘基可以插入另一個(gè)單體的疏水口袋。根據(jù)這種模式設(shè)計(jì)的一些肽類分子模擬配體已有相關(guān)報(bào)道,但還未有相關(guān)的配體分子被鑒定,可能是由于這個(gè)疏水口袋在S-S二聚化狀態(tài)期間,一直被單體自身或另一個(gè)單體的色氨酸殘基所占據(jù),進(jìn)而影響了對(duì)相關(guān)分子機(jī)制的更進(jìn)一步深入研究。在體外進(jìn)行S-S二聚化過(guò)程的抑制研究也許不是最好的方法,我們需要考慮其他方式,研究小分子在調(diào)節(jié)細(xì)胞黏附分子組裝方面的功能。
本研究中,我們對(duì)這個(gè)過(guò)程下的任何狀態(tài)進(jìn)行了小分子篩選,該篩選文庫(kù)包括類似藥物的小分片段,鑒定出了一種全新的可以特異性與P-鈣粘素蛋白結(jié)合的小分子。該化合物分子能夠通過(guò)調(diào)節(jié)X二聚化抑制細(xì)胞之間的黏附,間接的抑制了能夠穩(wěn)定X二聚物的疏水鍵的形成。
該化合物文庫(kù)包含1973種化合物。做為初步的篩選,進(jìn)行了分子之間相互作用檢測(cè)。P-鈣粘素蛋白的單體突變蛋白稱為REC12,其中精氨酸被插入到EC12的氨基端,通過(guò)生物素-鏈霉親和素捕獲法,被固定在生物傳感器芯片表面,含有100uM化合物的溶液流經(jīng)生物芯片表面。根據(jù)固定的分子水平,REC和化合物的分子量,推測(cè)結(jié)合應(yīng)答的RMAX在20RU左右。在1973種化合物文庫(kù)中。144種化合物的結(jié)合應(yīng)答效率超過(guò)10RU。
作為更進(jìn)一步的篩選過(guò)程,我們將一種稱為EC12的蛋白結(jié)構(gòu)狀態(tài)固定在CM5傳感器芯片表面。這種結(jié)構(gòu)是位于單體和S-S二聚體之間的一種平衡態(tài)。我們進(jìn)行了一種稱為ABA的檢測(cè)實(shí)驗(yàn),即A和B兩種溶液都流經(jīng)芯片表面。2uM的EC12被稱為溶液A,含有上述第一輪篩選后的化合物的溶液為溶液B,濃度為100uM.對(duì)于A溶液,單體結(jié)合應(yīng)答以及單體和芯片之間的共價(jià)結(jié)合應(yīng)答,都被檢測(cè)到;對(duì)于B溶液,兩種化合物明顯的抑制了結(jié)合應(yīng)答效率(請(qǐng)見(jiàn)Fig. 1C)。這些化合物破壞了二聚化的形成,使得單體不能共價(jià)結(jié)合到芯片表面。但只有其中一種化合物,Hit 1, 顯示出了劑量依賴性結(jié)合應(yīng)答。單體突變的KD值約為916uM(請(qǐng)見(jiàn)Fig. 1e)。
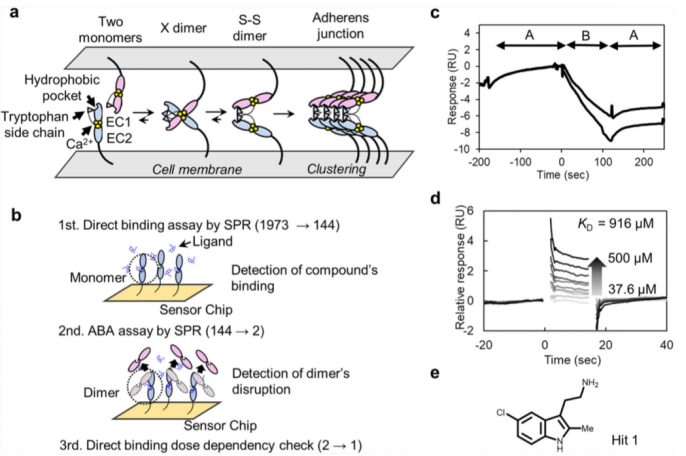
Fig. 1 SPR對(duì)潛在化合物篩選示意圖
我們進(jìn)行了X-射線晶體衍射實(shí)驗(yàn),用以鑒定Hit 1與單體,REC12的結(jié)合位點(diǎn),闡明Hit 1的抑制效應(yīng)。REC12經(jīng)結(jié)晶后,與終濃度10mM的Hit 1進(jìn)行結(jié)合,該復(fù)合物經(jīng)衍射后分辨率為2.30 ? (請(qǐng)見(jiàn)Fig. 2a, Table 1)。位于EC1和EC2之間的結(jié)構(gòu)域的淺腔內(nèi),觀察到了可以用于建模的Hit 1的電子密度分布。Hit 1一經(jīng)與REC12結(jié)合,相對(duì)于未結(jié)合Hit 1的REC12,其Y140側(cè)鏈發(fā)生了偏轉(zhuǎn)(請(qǐng)見(jiàn)Fig. 2b),Y140和Hit 1形成了一個(gè)CH–π的相互作用。這里需要注意,無(wú)論是結(jié)合Hit 1還是未結(jié)合Hit 1的REC12結(jié)構(gòu),都是高度的同型的,因此差異不應(yīng)來(lái)自高度同型的Hit 1時(shí),會(huì)有一個(gè)水分子位于REC12晶體結(jié)構(gòu)的腔內(nèi)。因此,與Hit 1結(jié)合的啟動(dòng)效應(yīng)有可能來(lái)自于和Y140殘基的互作以及脫水作用。同時(shí)發(fā)現(xiàn),Hit 1和R68, V98, T99, D100和D137存在范德華力引起的相互作用(請(qǐng)見(jiàn)Fig. 2C)。為了更進(jìn)一步確認(rèn)在Hit 1結(jié)合后的晶體結(jié)構(gòu)中觀察到的電子密度,我們通過(guò)SPR相關(guān)技術(shù),檢測(cè)了REC12 Y140R和Hit 1的結(jié)合效應(yīng),發(fā)現(xiàn)Hit 1未能夠和該突變結(jié)合,證實(shí)了Y140位點(diǎn)在相互作用中的作用。在結(jié)合口袋周圍的數(shù)個(gè)氨基酸殘基,比如Y140,D100和Q101經(jīng)報(bào)道對(duì)于X二聚化至關(guān)重要。在X二聚體中,可觀測(cè)到Y(jié)140和K14,Q101和D100之間存在氫鍵。盡管Hit 1未能直接破壞這些氨基酸殘基之間的氫鍵,但是該區(qū)域結(jié)構(gòu)的改變顯著抑制了X的二聚化。
有趣的是,當(dāng)我們將P-鈣粘素-Hit 1復(fù)合物結(jié)構(gòu)與E-鈣粘素結(jié)構(gòu)進(jìn)行疊加時(shí),我們發(fā)現(xiàn)E-鈣粘素的腔室體積小于P-鈣粘素的腔室體積。通過(guò)CASTp 30對(duì)P-鈣粘素,E-鈣粘素結(jié)構(gòu)和P-鈣粘素單體-Hit 1復(fù)合物結(jié)構(gòu)的腔室體積進(jìn)行測(cè)量,也證明了這一點(diǎn),分別是12.2 ? 3 , 5.4 ? 3 , 21.4 ? 3。該結(jié)果表明較之于E-鈣粘素,P-鈣粘素的腔室更適合于結(jié)合Hit 1分子。此外,F(xiàn)TPap分析得到的31個(gè)結(jié)合區(qū)域,都存在于P-鈣粘素中。SPR數(shù)據(jù)同樣表明,Hit 1不與E-鈣粘素結(jié)合,但是能夠與E-鈣粘素N140Y突變體結(jié)合(請(qǐng)見(jiàn)Fig. 2e)。因此,與Hit 1的結(jié)合腔室,能夠做為P-鈣粘素選擇性配體設(shè)計(jì)的起始點(diǎn)。
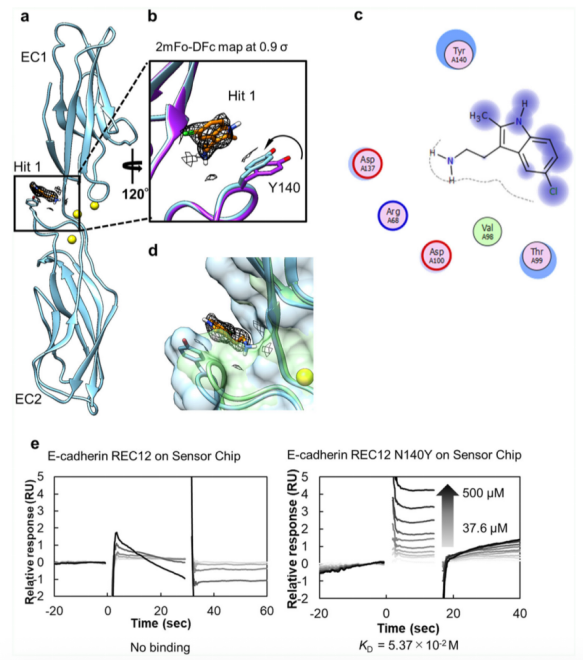
Fig. 2 Hit 1與P-鈣粘素蛋白結(jié)合位點(diǎn)鑒定
為了檢測(cè)Hit 1對(duì)X二聚化的影響,本文利用了氫-氘交換質(zhì)譜分析法(HDX-MS)。該方法能夠檢測(cè)在Hit 1存在與否的情況下,X二聚物的互作面有多大程度暴露在溶劑中。在采用 LC-MS 進(jìn)行檢測(cè)之前先進(jìn)行胃蛋白酶處理,從而能夠從 P-鈣黏蛋白分子的幾乎所有區(qū)域分離出肽片段。在Hit 1存在與否的情況下,檢測(cè)每一段肽段的HDX比率(氫氘交換比率)。在相互作用界面之一(殘基134-140和137-147)的相應(yīng)區(qū)域,該區(qū)域由于在EC12的N端插入了甲硫氨酸,能夠形成X二聚物,但不能形成S-S二聚物,因此抑制了strand-swapping,Hit 1存在時(shí)的氫氘交換比率高于未結(jié)合Hit 1時(shí)(請(qǐng)見(jiàn)Fig. 3a)。該結(jié)果表明,Hit 1對(duì)X二聚物的互作界面能夠產(chǎn)生影響。
為了進(jìn)一步研究Hit 1如何影響X二聚化,X二聚物(MEC12)經(jīng)結(jié)晶后,與終濃度10mM的Hit 1進(jìn)行結(jié)合,形成的復(fù)合物結(jié)構(gòu)的分辨率為2.45 ?(請(qǐng)見(jiàn)Fig. 3b)。與上述發(fā)現(xiàn)Hit 1的過(guò)程中相同,能夠檢測(cè)到三個(gè)獨(dú)立的電子密度分布,其中的兩個(gè)位于Y140殘基周圍的腔室,如Fig. 2a中所示。另一個(gè)位于EC2結(jié)構(gòu)域的交匯處(請(qǐng)見(jiàn)Fig. 3b)。位于腔內(nèi)的這些 Hit 1 分子與 Y140 形成了π-π相互作用,并與口袋周圍的殘基或水分子形成了氫鍵連接。由于Hit 1分子能夠與X二聚物的腔室結(jié)合,與口袋周圍的殘基形成多種非共價(jià)相互作用,Hit 1和X二聚物結(jié)合的親和性似乎更高。該推測(cè)經(jīng)switchSENSE技術(shù)進(jìn)行了確認(rèn)。Hit 1分子與EC2結(jié)構(gòu)域交匯處的互作主要產(chǎn)生于范德華力。X二聚化中的Y140周圍的腔室結(jié)構(gòu)與單體的不同,Hit 1的結(jié)合模式較單體也有可能存在差異。
與X二聚物無(wú)輔因子的狀態(tài)相比,與Hit 1結(jié)合的X二聚物呈現(xiàn)出較顯著的結(jié)構(gòu)變化。EC1-EC2結(jié)構(gòu)域角度變得平滑,B鏈相對(duì)于A鏈發(fā)生了轉(zhuǎn)變(請(qǐng)見(jiàn)Fig. 3c)。該角度的結(jié)構(gòu)變化推測(cè)可能是由于兩個(gè)Hit 1分子結(jié)合在Y140周圍的腔室引起。在 X 多聚體的界面處,K14 的側(cè)鏈與 A138 的主鏈之間形成的氫鍵(即在 Hit 1 結(jié)合時(shí)形成的那一個(gè))被破壞了(請(qǐng)見(jiàn)Fig. 3d),可能導(dǎo)致了兩個(gè)單體分子的位置轉(zhuǎn)變。
為了進(jìn)一步確認(rèn)K14側(cè)鏈和A138 的主鏈之間形成的氫鍵對(duì)X二聚化形成的重要性,本文使用了尺寸排阻色譜 - 多角度光散射,分別測(cè)量了X二聚物(MEC12)和X二聚物突變體K14A的分子量。得到的分子量大小分別是X二聚物為33.8kDa, X二聚物K14A突變體為24.2kDa。
由于在Hit 1結(jié)合的X二聚物中,K14和側(cè)鏈A138之間的關(guān)鍵互作被破壞,我們推測(cè)X二聚物的結(jié)構(gòu)可能代表著X二聚化形成過(guò)程中的一種不穩(wěn)定狀態(tài)。為了檢測(cè)無(wú)輔因子存在時(shí)的X二聚物的不穩(wěn)狀態(tài)或溶液狀態(tài)下的動(dòng)力學(xué),我們進(jìn)行了分子動(dòng)力學(xué)的模擬。進(jìn)行了三次各持續(xù) 100 納秒的獨(dú)立模擬,并通過(guò) Cα 原子的均方根偏差(RMSD)來(lái)確認(rèn)軌跡的收斂性。由此我們計(jì)分別計(jì)算出了K14A-A138之間我們分別計(jì)算的供體氫鍵和受體氧分子之間的距離(請(qǐng)見(jiàn)Fig. 3f),數(shù)據(jù)表明在頻率分布上存在兩個(gè)峰,一個(gè)是 2.6 ?,另一個(gè)是4.8 ?。這種非對(duì)稱特性在所有的軌跡中被檢測(cè)到。在Hit 1分子結(jié)合下的蛋白晶體結(jié)構(gòu)中,距離則分別為 4.6 和2.6 ?。上述結(jié)果表明,在氫鍵形成時(shí),無(wú)輔因子狀態(tài)下的X二聚物動(dòng)力學(xué)可能存在兩種狀態(tài),與Hit 1結(jié)合狀態(tài)下的X二聚物的兩種晶體動(dòng)力學(xué)狀態(tài)一致。換句話說(shuō),apo狀態(tài)的 X 大分子能夠呈現(xiàn)出與在晶體結(jié)構(gòu)中與 Hit 1 結(jié)合的 X 大分子所呈現(xiàn)的構(gòu)象非常接近的構(gòu)象,即便沒(méi)有 Hit 1 化合物的存在也是如此。表明,在晶體結(jié)構(gòu)中觀測(cè)到的Hit 1結(jié)合下的X二聚物的狀態(tài)是X二聚化過(guò)程中的一種狀態(tài)。總的說(shuō)來(lái),Hit 1的結(jié)合可能會(huì)通過(guò)將X二聚物限定在二聚化過(guò)程的一種狀態(tài)下,進(jìn)而干擾對(duì)X二聚化非常重要的氫鍵的形成。和orthosteric抑制劑不同,Hit 1不會(huì)直接的阻止兩個(gè)單體之間氫鍵的形成,而是可能通過(guò)改變單體結(jié)構(gòu),進(jìn)而阻止氫鍵的形成。
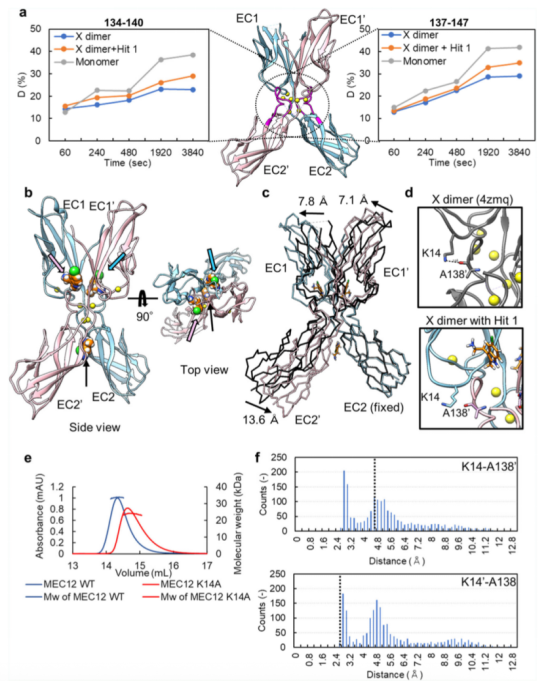
Fig. 3 Hit 1對(duì)蛋白二聚化的影響鑒定
我們進(jìn)一步通過(guò)細(xì)胞-聚集檢測(cè)技術(shù),驗(yàn)證了Hit 1是否能夠抑制細(xì)胞之間的黏附作用。我們通過(guò)Flp-In-CHO體系,構(gòu)建了一種表達(dá)P-鈣粘素的CHO細(xì)胞系。在該檢測(cè)中,除了P-鈣粘素蛋白,細(xì)胞外蛋白經(jīng)胰蛋白酶消化,使得細(xì)胞之間的黏附和細(xì)胞聚集體的形成僅僅取決于P鈣粘素分子的相互作用。實(shí)驗(yàn)表明,空白對(duì)照組CHO細(xì)胞之間未能夠形成大的細(xì)胞聚集體(請(qǐng)見(jiàn)Fig. 4a,c)。在胰蛋白酶消化后,培養(yǎng)基替換成不含鈣離子的培養(yǎng)基,以防止細(xì)胞聚集,通過(guò)添加1mM的CaCl2促進(jìn)細(xì)胞聚集。在細(xì)胞發(fā)生聚集反應(yīng)后,使用微流控成像(MFI)設(shè)備測(cè)量細(xì)胞聚集體尺寸大小分布。在MFI的測(cè)量中,大于40um的細(xì)胞聚集體被定義為P-鈣粘素依賴的細(xì)胞聚集,因?yàn)樵贓DTA存在時(shí),會(huì)有大量的大小在25-40um的細(xì)胞聚集體存在。為了對(duì)數(shù)量進(jìn)行標(biāo)準(zhǔn)化,通過(guò)計(jì)算大于40um-100um的顆粒數(shù)與大于25-100um的顆粒數(shù)的比值實(shí)現(xiàn)。在對(duì)照實(shí)驗(yàn)中,EDTA能夠抑制大的聚集體(40-100um)的形成,因此細(xì)胞黏附取決于鈣依賴性的P-鈣粘素分子的相互作用(請(qǐng)見(jiàn)Fig. 4a,c)。
當(dāng)同時(shí)添加Hit 1和1mM的CaCl 2分子時(shí),聚集體的形成呈現(xiàn)Hit 1濃度依賴的方式(請(qǐng)見(jiàn)Fig. 4b, c)。而當(dāng)Hit 1加入到已經(jīng)形成聚集體的培養(yǎng)體系中時(shí),在檢測(cè)時(shí)間段,形成的聚集體仍舊穩(wěn)定存在,而EDTA能夠部分的破壞已經(jīng)形成的聚集體。這些結(jié)果說(shuō)明,Hit 1是通過(guò)阻止X二聚化形成過(guò)程中兩個(gè)單體分子的互作,一旦細(xì)胞聚集體形成,Hit 1是無(wú)法破壞它們的互作穩(wěn)定性的。
為了對(duì)Hit 1在X二聚化過(guò)程中的效應(yīng)進(jìn)行量化分析,我們進(jìn)行了脂質(zhì)體聚集檢測(cè)。我們首先制備了帶有 C 端組氨酸標(biāo)簽的 X 二聚體(MEC12),然后將該蛋白質(zhì)與一種基于 DOPC 的脂質(zhì)體一起孵育,該脂質(zhì)體中含有 10% 的具有鎳吸附作用的脂質(zhì) DOGs-NTA-Ni,并在 EDTA 存在下使 P-鈣黏蛋白分子失活。在這些條件之下,在650nm的吸收光下,未觀測(cè)到聚集(請(qǐng)見(jiàn)Fig. 4d)。我們也同時(shí)觀測(cè)到,脂質(zhì)的聚集僅僅在MEC12和 CaCl 2都存在時(shí)發(fā)生(請(qǐng)見(jiàn)Fig. 4d)。一旦CaCl 2添加到溶液中,脂質(zhì)體聚集,推測(cè)是通過(guò)被捕獲在脂質(zhì)體表面的C端帶有組氨酸標(biāo)簽的X二聚物(MEC12)的二聚化引起,可通過(guò)逐漸增加的650nm處的吸光值檢測(cè),該吸光值最后可達(dá)到平臺(tái)期(請(qǐng)見(jiàn)Fig. 4d)。通過(guò)使用一相結(jié)合的指數(shù)衰減方程模型,計(jì)算出了脂質(zhì)體聚集的觀測(cè)速率常數(shù)(kobs),以此作為 X 雙聚體化動(dòng)力學(xué)的指標(biāo)。Hit 1存在時(shí)的速率常數(shù)(k obs = 0.0033) 顯著低于Hit 1未存在時(shí)的速率常數(shù) (k obs = 0.0086)(請(qǐng)見(jiàn)Fig. 4e)。此處需注意,這并非是由于鈣粘分子的激活引起,因?yàn)橥ㄟ^(guò)DSC測(cè)量可知,無(wú)論Hit 1存在與否,鈣離子與鈣粘素分子的結(jié)合并未被Hit 1所抑制。該結(jié)果表明,Hit 1通過(guò)間接的破壞對(duì)X二聚化形成關(guān)鍵的氫鍵,抑制了X二聚化的過(guò)程,進(jìn)而抑制了細(xì)胞的聚集。
我們利用兩種細(xì)胞系分析了Hit 1對(duì)于內(nèi)源性P-鈣粘素分子介導(dǎo)的細(xì)胞黏附的效應(yīng),以及對(duì)于不表達(dá)P-鈣粘素分子的細(xì)胞聚集效應(yīng)。其中HCT116表達(dá)內(nèi)源性P-鈣粘素分子,MCF7細(xì)胞不表達(dá)P-鈣粘素分子。將細(xì)胞種植在培養(yǎng)板中,并添加100uM的Hit 1分子,細(xì)胞聚集程度通過(guò)對(duì)整體區(qū)域面積下的聚集體進(jìn)行定量分析。由于HCT116和MCF7的每一個(gè)單細(xì)胞區(qū)域都不同,標(biāo)準(zhǔn)化后的細(xì)胞區(qū)域經(jīng)Hit 1存在時(shí),每一個(gè)濃度下的整體細(xì)胞區(qū)域比上Hit 1不存在時(shí)整體細(xì)胞區(qū)域計(jì)算得到。在100uM的Hit 1存在時(shí),HCT116細(xì)胞標(biāo)準(zhǔn)化后的細(xì)胞區(qū)域,下降到未加入Hit 1時(shí)的細(xì)胞區(qū)域的75.7%,但Hit 1對(duì)MCF7細(xì)胞標(biāo)準(zhǔn)化后的區(qū)域沒(méi)有產(chǎn)生影響(請(qǐng)見(jiàn)Fig. 4f)。已有研究報(bào)道,P-鈣粘素蛋白敲除后對(duì)腫瘤轉(zhuǎn)移具有一定的影響,以及對(duì)細(xì)胞內(nèi)信號(hào)通路的影響。本研究中結(jié)合已有的表型研究表明,在表達(dá)P-鈣粘素分子的腫瘤細(xì)胞中,Hit 1能夠抑制細(xì)胞之間的黏附作用,進(jìn)而導(dǎo)致β -catenin信號(hào)通路的下調(diào)及細(xì)胞凋亡信號(hào)的激活。
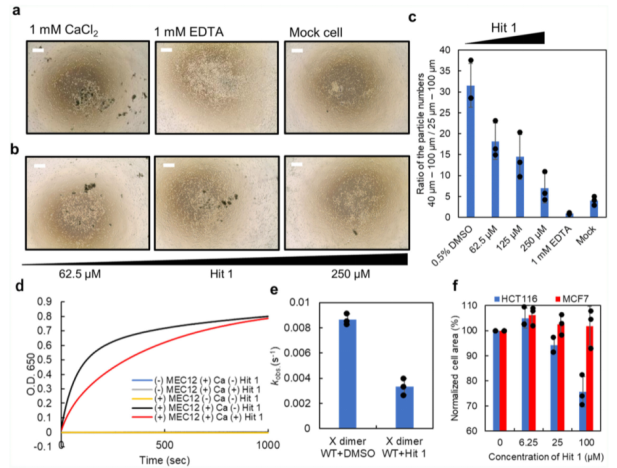
Fig. 4 Hit 1抑制細(xì)胞黏附檢測(cè)
由于Hit 1以及初篩鑒定的許多化合物都包括陽(yáng)離子功能基團(tuán),我們因此通過(guò)基于吲哚的商業(yè)化合物:tryptamine, tryptophan, 和 auxin 鑒定了陽(yáng)離子功能基團(tuán)對(duì)于結(jié)合活性的重要性(請(qǐng)見(jiàn)Fig. 5a)。令人驚訝的是,只有tryptophan與 P-鈣黏蛋白結(jié)合,這可能是由于Hit 1結(jié)合口袋周圍蛋白表面存在著負(fù)電荷。該結(jié)果表明,在進(jìn)一步的藥物發(fā)現(xiàn)中,不應(yīng)對(duì)陽(yáng)離子基團(tuán)進(jìn)行修飾改造。根據(jù)Hit 1和P-鈣粘素蛋白的2D相互作用圖譜的研究(請(qǐng)見(jiàn)Fig. 2c),我們制定了兩個(gè)進(jìn)一步合成的策略:(1)將吲哚環(huán)第二個(gè)位置的甲基替換成羧基,這將導(dǎo)致羧基和R68,氨基和D137之間形成鹽橋;(2)將吲哚環(huán)中 C5 位置的氯原子替換為一種更大分子量的官能團(tuán),抑制第二個(gè)單體的接近。我們首先合成了3-(2-氨基乙基)-5-苯基-1H-吲哚-2-羧酸(簡(jiǎn)稱 Hit 1-羧酸,請(qǐng)見(jiàn)Fig. 5b),但是從SPR親和性檢測(cè)來(lái)看,它與REC12的親和性并不優(yōu)于Hit 1,也不具備細(xì)胞聚集抑制效應(yīng)。接下來(lái),我們合成了 2-(2-甲基-5-苯基-1H-吲哚-3-基)乙烷-1-胺(苯基-Hit 1)(請(qǐng)見(jiàn)Fig. 5b)。通過(guò)SPR互作檢測(cè)分析,我們觀測(cè)到一種劑量依賴性應(yīng)答;其與REC結(jié)合的親和性與Hit 1相同。在脂質(zhì)體聚集檢測(cè)中,苯基-Hit 1相比較于Hit 1,具有更強(qiáng)的抑制活性。tryptamine, auxin 和 tryptophan對(duì)聚集效應(yīng)影響較小(請(qǐng)見(jiàn)Fig. 5c)。Tryptamine 不具有大的功能基團(tuán),這可能是它不能抑制脂質(zhì)體聚集的原因。在細(xì)胞聚集檢測(cè)中,苯基-Hit 1相較于Hit 1具有更強(qiáng)的抑制效應(yīng),且苯基-Hit 1與X二聚體的結(jié)合親和性要強(qiáng)于單體P-鈣粘素蛋白。
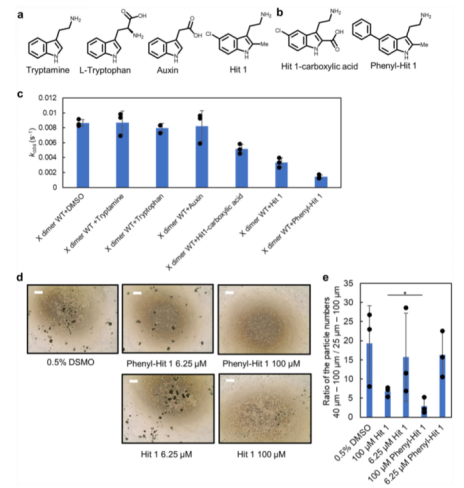
Fig. 5 抑制劑設(shè)計(jì)策略

